菊石長這個樣子,還真不知道它怎麼游泳呢!
北海道異常卷曲菊石之一,
在白堊紀末期發展到了巔峰,隨後而來的就是菊石大滅絕。
我的想像是因為沒有天敵的追殺,所以演化出不需要會游泳的異常卷曲的殼形,太過安逸,代價是大滅絕?
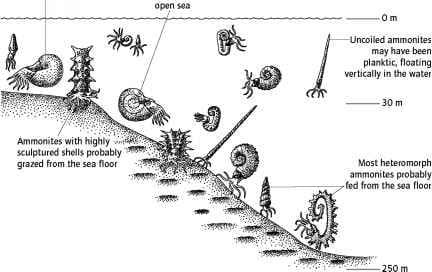
據研究資料顯示(如圖),有些菊石可能是底棲性進食,跟我們傳統認為的噴射後退及上下升降有很大的不同。
這件清修的很好,也好擺飾。
18-24-17




這一面是原石自然水洗風化掉

有上膠保固

論文參考資料:
Neale Monks和Jeremy R. Young
物品編號:1.1.1A
版權古生物學協會,1998年1月28日
提交:1997年5月27日。接受:1997年11月23日
前言
一般對菊石的功能形態分析,假設動物充滿了整個住室,並且動物的運動(例如將頭部和臂部撤回到殼體中)對方向幾乎沒有影響。
這裡提出了另一種可能性:菊石動物體積小,可移動,並且在受到威脅時能夠遠離住室口。
並分析了這種解剖學對勾菊石的流體靜力學的影響。該分析表明,身體的運動將顯著改變質量分佈,從而改變方向。
介紹
對化石頭足類動物的功能形態的解釋傾向於與現存的頭足類動物比較,但是這種的比較存在一個問題…我們不清楚鸚鵡螺與菊石的類似程度。
鸚鵡螺透過相關的肌肉,在受到威脅時,可以將頭部和觸手拉入殼體。
而住室洞口也由堅硬的罩密封,功能類似於腹足類鰓蓋。
但是,鸚鵡螺的體腔較短,長度約為螺旋的三分之一。相比之下,菊石的住室非常長並且通常很窄。此外,菊石缺乏罩或鰓蓋。[注意:現在人們普遍認為aptychi是頜骨的一部分(Lehmann 1981)]。
因此,假設菊石的軟體部分填充在殼體的整個住室中並不是必然正確的,並且可能沒有類似鰓蓋的方式,菊石可能已經能夠完全進入殼體。
異形菊石(勾菊石)中的住室是重要的,因為軟體(例如頭部,臂部,內臟)與殼體的相對應關聯是暸解這些動物如何生活和運作的關鍵部分。Trueman(1941)關於這個主題的討論被廣泛引用(例如,在無脊椎動物古生物學論文中,Arkell等人,1957)。
Trueman認為整個住室都被軟體填滿了,並用它來改變菊石的重心位置和浮力。基於該模型的菊石功能解釋的特徵在於單個穩定取向,其中質心垂直地位於浮力中心下方。
最近,Kakabadzé和Sharikadzé (1993)提出異形菊石具有兩個穩定的方向,一個具有向上的孔(如先前的解釋中),另一個具有朝向海底傾斜的孔。這種替代方向將使異形菊石能夠直接從海底進食。
Kakabadzé和Sharikadzé(1993)也提出,這些菊石可以通過在殼內移動體液來調節它們的浮力。這將改變質心的位置,從而改變殼的方向。
Kakabadzé和Sharikadzé(1993)模型假設菊石能夠快速改變殼內流體的分佈或數量。就目前所知,氣室內的流體運動緩慢。
在Nautilus,即使通過調整整體浮力來幫助晝夜遷移,室內流體量的變化也太慢(Chamberlain 1991)。墨魚具有高度改性和特徵性的殼,分為許多小室,由conchiolin牆細分,並展現出寬闊的虹膜區而不是狹窄的虹膜(Denton&Gilpin-Brown 1961a)。這似乎允許墨魚比Nautilus更快地改變殼內的浮力(Denton&Gilpin-Brown 1961b)。
即便如此,烏賊的浮力調節似乎不能用作上升或下沉的主要裝置,而是在達到優選深度時獲得中性浮力(Denton 1973)。
菊石的貝殼與烏賊的貝殼非常不同,因此更有可能是鸚鵡螺在這方面是一個合適的類別。
結果,問題仍然存在,雖然可能已經有可能改變方向,但沒有提出完全令人信服的機制來確定如何實現這一目標。我們提出了第三種可能的解剖學方法:異形菊石動物是一種相對較小的移動生物,能夠在其浮動殼內移動,而不像現代腹足動物。與之前的模型一樣,這允許方向的顯著變化,因為在這種情況下,當身體在殼體內移動時,質量分佈的變化。
肌肉疤痕提供了菊石中軟部分形態的唯一直接證據。它們始終位於生活室後部附近,靠近最後一個隔膜(Crick 1898)。這種肌肉發達的肌肉與我們的小型可伸縮身體模型兼容,但顯然它並不能證明這一點。異形態菊石殼的特徵在於偏離大多數其他菊石的規則平面形狀。
分析
水下物體的方向由其浮力中心和質心的位置決定。浮力中心實際上是殼體旋轉的樞軸點。由於質心在靜止時會直接位於此點之下,因此推斷其位置對於識別殼體方向至關重要。為了證明在自由浮動的殼體內移動小而重的物體的效果,因此需要定位浮力中心和質心。
浮力中心是體內浸沒的置換流體的重心。在Trueman(1941)和Kakabadzé&Sharikadzé(1983)提出的模型中,流動的水大約等於氣室和生活室的體積,因為兩者都被填充。相比之下,我們的模型減小了動物的大小,使得大部分生活室都是空的。
質心是從重力的角度來看,總質量可以被認為是集中的點。對於Trueman來說,菊石中最重的部分是住室,它充滿了大部分的活體。
由於我們對菊石軟體部分的密度或質量分佈或殼體內有多少流體一無所知,因此質心更難以推斷。我們遵循Trueman的假設,即它靠近身體的質心,這是動物中最重要的部分,它被認為具有均勻的密度。
討論
克林格(Klinger,1980)證明勾菊石必須是頭部向下的生物,並且表明這些動物可能在海底覓食,可能在受到干擾時垂直向上噴射。
Ebel(1992)重建了許多異形菊石(如Scaphites和Nipponites))大體不能縮回殼體,總體上它們是負浮力的(浮子與身體成比例太小)。相反,Ebel(1992)提出這些動物像現代章魚一樣在肌肉臂上沿著海底爬行。
我們的模型與Ebel(1992)提出的建議形成鮮明對比,他提出,與鸚鵡螺身體的大小遠遠不同,它更小,更像是腹足類動物。然而,像Ebel(1992)和Klinger(1980)一樣,我們相信許多異形菊石是底棲生物。為了支持這種解釋,有充分證據表明至少有一些菊石存在於海床上。腸道內容顯示,一些菊石以底棲魚類,介形蟲,海百合,甲殼類和其他菊石為食,也可能是食腐生物(Lehmann 1973,Lebrun 1996)。
在大多數菊石(和所有異形體)中沒有一個噴射裝置,如果存在的話,是弱發展的,並且在鸚鵡螺看到了那種可操縱的噴射不太可能。
事實上,Géczy(1960)認為菊石可能缺乏游泳能力。侏羅紀和白堊紀的異形菊石通常在粘土或泥灰岩而不是砂質相中發現,這也表明異形菊石是基質依賴的(Marcinowski&Wiedmann 1990)
有些例子似乎是作為底棲生物的一部分保存的菊石樣本“嵌套”在Eupachydiscus的殼內(Matsumoto&Nihongo 1979)。
最後,注意到在底層水域缺氧期間一些異形菊石的周期性消失(例如,Scaphites和Hamites)來自Western Interior sea,Batt 1989)。此外,可以注意到Hamitidae基本上局限於陸架沉積物(Westermann 1996)。
在長體腔內有一個小的移動體,可以看到異形菊石,就像一個帶有移動洞穴或洞穴的小章魚。在覓食的時候,住室口將朝向沉積物傾斜,允許動物覓食沉積物,可能將其自身與其手臂一起拉動,挖掘小型軟體動物,蠕蟲和甲殼類動物。
當遇到危險時,菊石會深深地進入殼內,並等待危險通過。由於重心位置的變化,殼體將旋轉離開。菊石可能依賴於外殼的機械防禦(使殼難以壓碎)和肋骨(以阻止剝落),而不是遊走。菊石的長體室也會使動物難以拔出。類似的適應性在腹足動物中有很好的記載(Vermeij 1993)。
這種生活方式類似於Kakabadzé和Sharikadzé(1993)所建議的生活方式,但由於殼體相對於氣室的取向變成了動物在殼體內的位置和殼體本身的幾何形狀的函數; 消除了對殼體周圍流動的要求。
對異形態菊石形態的其他解釋包括從殼體中部分出現長住室腔(Jacobs和Landmann 1993); 甚至是一種自由生活的菊石動物,使用殼作為蛋殼(Lewy,1996)。
然而,對於所有這些解釋,證據充其量只是間接的。我們的模型的優點是在行為或解剖學方面要求與現代頭足類動物的較少分歧,符合對菊石生態學更為保守的觀點,並暗示迄今為止未考慮的生活方式的可能性。
結論
異形菊石的取向取決於動物的軟體部分在殼內的定位。我們認為身體是一個相對較小,緊湊的質量,只是部分填充了生活室。當身體從洞口移動到最後的隔膜然後再返回時,預計質心將以使殼體旋轉的方式移動。這種變化的大小是殼形的函數,尤其是捲曲模式。該效果可以提供異形殼幾何形狀的功能性解釋。此外,該模型的更廣泛應用可能導致對其他菊石群的生命模式的重新解釋。

 留言列表
留言列表